
Seymouria -Seymouria
|
Seymouria |
|
|---|---|

|
|
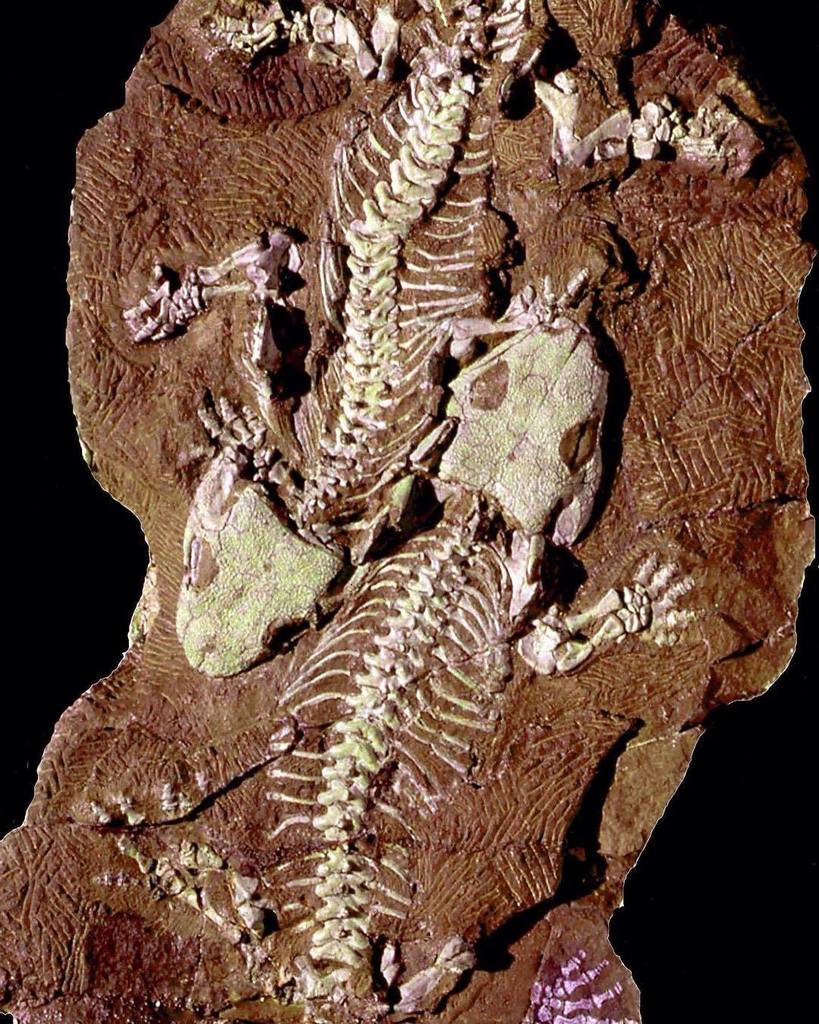
| Un fossile de Seymouria baylorensis exposé au Muséum national d'histoire naturelle | |
|
Classement scientifique |
|
| Royaume: | Animalia |
| Phylum: | Accords |
| Commander: | † seymouriamorpha |
| Famille: | † Seymouriidae |
| Genre: |
† Seymouria Broili, 1904 |
| Espèce | |
|
|
| Synonymes | |
|
|
Seymouria est un genre éteint de Seymouriamorph du Permien inférieur d' Amérique du Nord et d' Europe . Bien qu'ils soient des amphibiens (au sens biologique), les Seymouria étaient bien adaptés à la vie sur terre, avec de nombreusescaractéristiques reptiliennes , si nombreuses, en fait, qu'ona d'abord penséque Seymouria était un reptile primitif. Il est principalement connu à partir de deux espèces, Seymouria baylorensis et Seymouria sanjuanensis . L'espèce type, S. baylorensis , est plus robuste et spécialisée, bien que ses fossiles n'aient été trouvés qu'au Texas . En revanche, Seymouria sanjuanensis est plus abondant et plus répandu. Cette espèce plus petite est connue à partir de plusieurs fossiles bien conservés, dont un bloc de six squelettes trouvés dans la formation de Cutler au Nouveau-Mexique et une paire de squelettes entièrement développés de la formation de Tambach en Allemagne , qui ont été fossilisés l'un à côté de l'autre.
Pendant la première moitié du 20e siècle, Seymouria était considéré comme l'un des reptiles les plus anciens et les plus « primitifs » connus. Les paléontologues ont noté à quel point la forme générale du corps ressemblait à celle des premiers reptiles tels que les captorhinidés , et que certaines adaptations des membres, de la hanche et du crâne étaient également similaires à celles des premiers reptiles, plutôt qu'à toute espèce d'amphibiens modernes ou éteints connus à l'époque. . Les membres et la colonne vertébrale fortement construits soutenaient également l'idée que Seymouria était principalement terrestre, passant très peu de temps dans l'eau. Cependant, dans les années 1950, des têtards fossilisés ont été découverts chez Discosauriscus , qui était un proche parent de Seymouria dans le groupe Seymouriamorpha . Cela montre que les seymouriamorphes (y compris Seymouria ) avaient un stade larvaire qui vivait dans l'eau, faisant ainsi de Seymouria non pas un vrai reptile, mais plutôt un amphibien qui était étroitement lié aux reptiles. Bien qu'il ne soit plus considéré comme le reptile le plus primitif, Seymouria est toujours un important fossile de transition documentant l'acquisition de caractéristiques squelettiques de type reptile avant l'évolution de l'œuf amniotique, qui caractérise les amniotes (reptiles, mammifères et oiseaux ).
Histoire

Des fossiles de Seymouria ont été trouvés pour la première fois près de la ville de Seymour , dans le comté de Baylor , au Texas (d'où le nom de l'espèce type, Seymouria baylorensis , faisant référence à la fois à la ville et au comté). Les premiers fossiles découverts étaient un groupe d'individus collectés par CH Sternberg en 1882. Cependant, ces fossiles ne seront pas correctement préparés et identifiés comme Seymouria avant 1930.
Divers paléontologues du monde entier ont récupéré leurs propres fossiles de Seymouria baylorensis à la fin du XIXe siècle et au début du XXe siècle. Les premiers fossiles explicitement nommés Seymouria étaient une paire de crânes incomplets, dont l'un était conservé avec quelques éléments pectoraux et vertébraux. Ces fossiles ont été décrits par le paléontologue allemand Ferdinand Broili en 1904, et sont maintenant conservés à Munich . Le paléontologue américain SW Williston a décrit plus tard un squelette presque complet en 1911 et a noté que « Desmospondylus anomalus », un taxon qu'il avait récemment nommé à partir de membres et de vertèbres fragmentaires, représentait probablement des individus juvéniles ou même embryonnaires de Seymouria .
De même, le paléontologue anglais DMS Watson a noté en 1918 que Conodectes , un genre douteux nommé par le célèbre Edward Drinker Cope en 1896, était probablement synonyme de Seymouria . Robert Broom (1922) a fait valoir que le genre devrait être appelé Conodectes puisque ce nom a été publié en premier, mais Alfred Romer (1928) s'y est opposé, notant que le nom Seymouria était trop populaire au sein de la communauté scientifique pour être remplacé. Pendant ce temps, Seymouria était généralement considéré comme un reptile très ancien, faisant partie d'un grade évolutif connu sous le nom de « cotylosaures », qui comprenait également de nombreux autres reptiles du Permien au corps robuste ou des tétrapodes ressemblant à des reptiles. Cependant, de nombreux paléontologues étaient incertains de son allégeance avec les reptiles, notant de nombreuses similitudes avec les embolomères , qui étaient incontestablement des amphibiens "labyrinthodontes". Cette combinaison de caractéristiques des reptiles (c'est-à-dire d'autres "cotylosaures") et des amphibiens (c'est-à-dire des embolomères) était la preuve que Seymouria était au centre de la transition évolutive entre les deux groupes. Quoi qu'il en soit, on n'en savait pas assez sur sa biologie pour conclure à quel groupe il faisait vraiment partie. Broom (1922) et le paléontologue russe Peter Sushkin (1925) ont soutenu un placement parmi les amphibiens, mais la plupart des études à cette époque le considéraient provisoirement comme un reptile extrêmement « primitif » ; ceux-ci comprenaient une redescription complète du matériel faisant référence à l'espèce, publiée par Theodore E. White en 1939.

Cependant, des preuves indirectes que Seymouria n'était pas biologiquement reptilien ont commencé à émerger. Dans les années 1940, plusieurs genres étaient liés à Seymouria dans le cadre du groupe Seymouriamorpha . Certains seymouriamorphes, tels que Kotlassia , présentaient des preuves d'habitudes aquatiques, et même Seymouria lui-même avait parfois été avancé pour posséder des lignes latérales, des structures sensorielles utilisables uniquement sous l'eau. Watson (1942) et Romer (1947) ont chacun renversé leur position sur la classification de Seymouria , le plaçant parmi les amphibiens plutôt que parmi les reptiles. La preuve la plus accablante est peut-être venue en 1952, lorsque le paléontologue tchèque Zdeněk Špinar a signalé des branchies conservées dans des fossiles juvéniles du seymouriamorph Discosauriscus . Cela prouvait sans équivoque que les seymouriamorphes avaient un stade larvaire et étaient donc des amphibiens, biologiquement parlant. Néanmoins, le grand nombre de similitudes entre les Seymouria et les reptiles soutenait l'idée que les seymouriamorphes étaient plutôt proches de l'ascendance des amniotes .

En 1966, Peter Paul Vaughn a décrit les restes de Seymouria de l' Organ Rock Shale de l' Utah . Ces restes, qui étaient un assortiment de crânes, représentaient une nouvelle espèce, Seymouria sanjuanensis . Les fossiles de cette espèce se sont avérés plus abondants et plus répandus que ceux de Seymouria baylorensis . Plusieurs autres espèces ont été nommées plus tard par Paul E. Olson , bien que leur validité ait été plus discutable que celle de S. sanjuanensis . Par exemple, Seymouria agilis (Olson, 1980), connu à partir d'un squelette presque complet de la Formation Chickasha de l' Oklahoma , a été réaffecté par Michel Laurin et Robert R. Reisz au parareptile Macroleter en 2001. Seymouria grandis , décrit un an plus tôt à partir d'un boîte crânienne trouvée au Texas, n'a été référée à aucun autre tétrapode, mais elle reste mal connue. Langston (1963) a signalé un fémur impossible à distinguer de celui de S. baylorensis dans les sédiments du Permien à l' Île-du-Prince-Édouard sur la côte est du Canada . Des restes squelettiques de type Seymouria sont également connus de la carrière Richards Spur en Oklahoma, comme décrit pour la première fois par Sullivan et Reisz (1999).
Un bloc de sédiments contenant six squelettes de S. sanjuanensis a été trouvé dans la formation Cutler du Nouveau-Mexique , comme décrit par Berman, Reisz et Eberth (1987). En 1993, Berman & Martens ont signalé les premiers restes de Seymouria en dehors de l'Amérique du Nord , lorsqu'ils ont décrit des fossiles de S. sanjuanensis de la formation de Tambach en Allemagne . La Formation de Tambach a également produit des fossiles de S. sanjuanensi d'une qualité similaire à ceux de la Formation de Cutler. Par exemple, en 2000, Berman et ses collègues ont décrit les "Tambach Lovers", deux squelettes complets et entièrement articulés de S. sanjuanensis fossilisés allongés l'un à côté de l'autre (bien qu'il ne soit pas possible de déterminer s'il s'agissait d'un couple tué lors de l'accouplement). La formation de Tambach a également produit les fossiles connus de Seymouria les plus jeunes du point de vue du développement , ce qui facilite les comparaisons avec Discosauriscus , qui est connu principalement à partir de juvéniles.
La description
Les individus Seymouria étaient des animaux robustes, avec une grosse tête, un cou court, des membres trapus et des pieds larges. Ils étaient assez petits, seulement environ 2 pieds (60 cm) de long. Le crâne était carré et à peu près triangulaire vu d'en haut, mais il était plus bas et plus long que celui de la plupart des autres seymouriamorphes. Les vertèbres avaient des arcs neuraux larges et gonflés (la partie au-dessus de la moelle épinière). Dans l'ensemble, la forme du corps était similaire à celle des reptiles contemporains et des tétrapodes ressemblant à des reptiles tels que les captorhinidés , les diadectomorphes et les parareptiles . Collectivement, ces types d'animaux ont été appelés « cotylosaures » dans le passé, bien qu'ils ne forment pas un clade (un groupe naturel basé sur les relations).
Le crâne

Le crâne était composé de nombreux petits os en forme de plaque. La configuration des os du crâne présents à Seymouria était très similaire à celle de tétrapodes et de parents tétrapodes bien plus anciens. Par exemple, il conserve un os intertemporel , qui est la condition plésiomorphe ("primitive") présente chez les animaux comme Ventastega et les embolomères . Les os du crâne étaient fortement texturés, comme c'était le cas pour les anciens amphibiens et les reptiles captorhinidés . De plus, la partie arrière du crâne avait une grande incision s'étendant le long de son côté. Cette incision a été appelée une encoche otique , et une incision similaire dans la même zone générale est commune à la plupart des amphibiens du Paléozoïque ("labyrinthodontes", comme on les appelle parfois), mais inconnue chez les amniotes. Le bord inférieur de l'encoche otique a été formé par le Squamosal , tandis que le bord supérieur tourné vers le bas a été formée par des rebords des supratemporal et tabulaires os (appelés brides otiques). Le tabulaire présente également un second rebord rabattu visible depuis l'arrière du crâne ; cette bride (appelée bride occipitale) reliée à la boîte crânienne et obscurcissait partiellement l'espace entre la boîte crânienne et le côté du crâne. Le développement des rebords otiques et occipitaux est plus important chez Seymouria (en particulier S. baylorensis ) que chez tout autre seymouriamorphe.
L'appareil sensoriel du crâne mérite également une mention pour un éventail de caractéristiques uniques. Les orbites ( orbites oculaires) étaient à peu près à mi-chemin de la longueur du crâne, bien qu'elles soient un peu plus proches du museau chez les juvéniles. Ils étaient plus rhomboïdaux que les orbites circulaires des autres seymouriamorphes, avec un bord antérieur aigu. Plusieurs auteurs ont noté que quelques spécimens de Seymouria possédaient des rainures indistinctes présentes dans les os entourant les orbites et devant l'encoche otique. Ces rainures étaient probablement des vestiges d'un système de lignes latérales , un réseau d'organes de détection de pression utiles pour les animaux aquatiques, y compris le stade larvaire présumé de Seymouria . De nombreux spécimens ne conservent aucun vestige de leurs lignes latérales, pas même les juvéniles. Environ au milieu des os pariétaux se trouvait un petit trou appelé foramen pinéal, qui aurait pu contenir un organe sensoriel appelé œil pariétal . Le foramen pinéal est plus petit chez Seymouria que chez les autres seymouriamorphes.
L' étrier , un os en forme de tige qui se situe entre la boîte crânienne et la paroi du crâne, était effilé. Il reliait la boîte crânienne au bord supérieur de l'encoche otique et servait probablement de conduit de vibrations reçues par un tympan (tympan) qui se trouvait vraisemblablement dans l'encoche otique. De cette façon, il pourrait transmettre le son du monde extérieur au cerveau. La configuration de l'étrier est intermédiaire entre les tétrapodes non amniotes et les amniotes. D'une part, sa connexion à l'encoche otique est inhabituelle, car les vrais reptiles et autres amniotes ont perdu une encoche otique, forçant le tympan et l'étrier à se déplacer vers le bas vers l' os carré de l'articulation de la mâchoire. D'autre part, la structure fine et sensible de l'étrier de Seymouria est une spécialisation par rapport à la plupart des tétrapodes non amniotes, qui ont un étrier épais inutile pour l'audition mais utile pour renforcer le crâne. L'oreille interne de Seymouria baylorensis conserve un évidement cochléaire situé derrière (plutôt qu'en dessous) du vestibule , et son canal semi-circulaire antérieur était probablement entouré d'un supraoccipital cartilagineux (plutôt qu'osseux) . Ces caractéristiques sont plus primitives que celles des vrais reptiles et synapsides.
Le palais (toit de la bouche) présentait certaines similitudes avec les tétrapodes amniotes et non amniotes. D'une part, il gardait quelques crocs isolés avec un repliement de l'émail interne en forme de labyrinthe, caractéristique des amphibiens "labyrinthodontes". D'autre part, les os du vomer à l'avant de la bouche étaient assez étroits et les choanes adjacentes (trous menant de la cavité nasale à la bouche) étaient grandes et rapprochées, comme dans les amniotes. Le palais est généralement constitué d'os solide, avec seulement des vacuités interptéryoïdes vestigiales (une paire de trous le long de la ligne médiane) séparées par un processus cultriforme long et mince (la branche avant de la base de la boîte crânienne). Outre les crocs isolés, le palais est également recouvert de petits denticules rayonnant à partir de la partie arrière des os ptérygoïdiens . Seymouria a quelques caractéristiques de type amniote du palais, telles que la présence d'une branche en forme de dent du ptérygoïde (anciennement connue sous le nom de bride transversale) ainsi qu'un os épiptérygoïde qui est séparé du ptérygoïde. Cependant, ces caractéristiques ont été observées chez divers tétrapodes non amniotes, elles ne signifient donc pas son statut d'amniote.
La mâchoire inférieure a conservé quelques caractéristiques plésiomorphes. Par exemple, le bord interne de la mandibule possédait trois os coronoïdes. La mandibule a également conservé au moins un grand trou le long de son bord interne connu sous le nom de fenêtre meckélienne, bien que cette caractéristique n'ait été confirmée que lors d'une nouvelle enquête en 2005 sur l'un des spécimens de la formation Cutler. Aucun de ces traits n'est commun chez les amniotes. La boîte crânienne avait une mosaïque de caractéristiques en commun avec divers tétrapodomorphes. Le système de rainures et d'ouvertures nerveuses sur le côté de la boîte crânienne était inhabituellement similaire à ceux du poisson Megalichthys , et la base cartilagineuse est une autre caractéristique plésiomorphe. Cependant, les artères carotides internes perforent la boîte crânienne près de l'arrière du complexe osseux, une caractéristique dérivée similaire aux amniotes.
Squelette post-crânien

La colonne vertébrale est assez courte, avec un total de 24 vertèbres devant la hanche. Les vertèbres sont gastrocentriques, ce qui signifie que chaque vertèbre a un composant plus gros, en forme de bobine, appelé pleurocentrum, et un composant plus petit en forme de coin (ou en forme de croissant depuis l'avant) appelé intercentrum. Les arcs neuraux, qui se trouvent au-dessus du pleurocentra, sont gonflés en de larges structures avec des zygapophyses en forme de table (plaques articulaires) environ trois fois plus larges que le pleurocentrum lui-même. Certaines vertèbres ont des épines neurales partiellement subdivisées au milieu, tandis que d'autres ont une section transversale de forme ovale. Les côtes des vertèbres dorsales s'étendent horizontalement et s'attachent aux vertèbres à deux endroits : l'intercentrum et le côté de l'arc neural. Le cou est pratiquement absent, seulement quelques vertèbres longues. La première vertèbre cervicale, l' atlas , avait un petit intercentrum ainsi qu'un pleurocentrum réduit qui n'était présent que chez les individus matures. Bien que le pleurocentrum atlantal (lorsqu'il est présent) était coincé entre l'intercentrum de l'atlas et l'intercentrum de la vertèbre axiale suivante (comme dans les amniotes), le faible développement osseux dans cette zone du cou contraste avec le complexe caractéristique de l'atlas-axe des amniotes. De plus, des études ultérieures ont révélé que l'atlas intercentrum était divisé en une partie gauche et une partie droite, ressemblant davantage à celle des tétrapodes de qualité amphibie. Contrairement à presque tous les autres tétrapodes paléozoïques (amniotes ou autres), Seymouria manque complètement de restes osseux d'écailles ou d'écailles, pas même les fines écailles du ventre circulaires des autres seymouriamorphes.
La ceinture pectorale (épaule) a plusieurs caractéristiques ressemblant à des reptiles. Par exemple, l' omoplate et la coracoïde (plaques osseuses situées au-dessus et au-dessous de l'orbite de l'épaule, respectivement) sont des os séparés, plutôt qu'une seule grande omoplate. De même, l' interclavicule était plat et en forme de champignon, avec une "tige" longue et fine. L' humérus (os de l'avant-bras) avait la forme d'un L légèrement tordu et carré, avec de grandes zones pour la fixation musculaire. Cette forme, qui a été qualifiée de « tétraédrique », est plésiomorphe pour les tétrapodes. D'autre part, l'humérus a également une adaptation de type reptile : un trou connu sous le nom de foramen entépicondylaire . Le rayon était à peu près en forme de sablier. Le cubitus est similaire, mais plus long en raison de la possession d'un processus olécrânien prononcé , comme cela est courant chez les tétrapodes terrestres mais rare chez les amphibies ou aquatiques. Le carpe (poignet) a dix os, et la main a cinq doigts robustes. Les os du carpe sont pleinement développés et se touchent étroitement, une autre indication de la territude. Les phalanges (os des doigts) diminuent de taille vers le bout des doigts, où elles se terminent chacune par un petit segment arrondi, sans griffe. La formule phalangienne (nombre de phalanges par doigt, du pouce au petit doigt) est 2-3-4-4-3.
Deux vertèbres sacrées (hanche) étaient présentes, bien que seule la première possédait une grosse côte robuste qui était en contact avec l'ilion (lame supérieure de la hanche). Certaines études ont soutenu qu'il n'y avait qu'une seule vertèbre sacrée, la deuxième sacrée supposée étant en fait la première caudale en raison d'une côte plus courte et plus incurvée que la première sacrée. Chaque ilion est bas et en forme de larme lorsqu'il est vu de côté, tandis que le dessous de la hanche dans son ensemble est formé par une seule plaque puboischiadique robuste, qui est rectangulaire lorsqu'elle est vue de dessous. Les orbites de la hanche et de l'épaule étaient dirigées à 45 degrés au-dessous de l'horizontale. Le fémur est aussi gros que l'humérus, et le tibia et le péroné sont des os robustes en forme de sablier similaires au radius et au cubitus. Le tarse (cheville) comprend 11 os, intermédiaires entre les tétrapodes antérieurs (qui en ont 12) et les amniotes (qui en ont 8 ou moins). Les pieds à cinq doigts sont assez similaires aux mains, avec la formule phalangienne 2-3-4-5-3.
Il n'y avait qu'une vingtaine de vertèbres caudales (queue) au maximum. Après la base de la queue, les caudales commencent à acquérir des épines osseuses le long de leur face inférieure, appelées chevrons . Ceux-ci commencent à apparaître à proximité de la troisième à la sixième caudale, selon le spécimen. Les côtes ne sont présentes que dans les cinq ou six premières caudales; ils sont longs à la base de la queue mais diminuent peu de temps après et disparaissent généralement autour de la même zone où apparaissent les chevrons.
Différences entre les espèces
Seymouria baylorensis et Seymouria sanjuanensis peuvent être distingués les uns des autres sur la base de plusieurs différences dans la forme et les connexions entre les différents os du crâne. Par exemple, la collerette osseuse renversée au-dessus de l'encoche otique (parfois appelée « corne tabulaire » ou « processus otique ») est beaucoup plus développée chez S. baylorensis que chez S. sanjuanensis ; il acquiert une forme triangulaire (vu de côté) à mesure qu'il s'étend vers le bas plus largement vers l'arrière du crâne. Chez S. sanjuanensis , l' os postfrontal entre en contact avec l' os pariétal au moyen d'une suture obtuse en forme de coin, tandis que la connexion entre les deux os est complètement droite chez S. baylorensis .

Certains auteurs ont avancé que les postpariétaux de S. baylorensis étaient plus petits que ceux de S. sanjuanensis , mais certains spécimens de S. sanjuanensis (par exemple, les "amoureux de Tambach") avaient également de petits postpariétaux. De plus, les "amoureux de Tambach" ont un os quadratojugal qui ressemble plus à celui de S. baylorensis qu'à celui de S. sanjuanensis . La combinaison des caractéristiques des deux espèces dans ces spécimens peut indiquer que les deux espèces font partie d'une lignée continue, plutôt que de deux chemins évolutifs fortement différenciés. De même, certaines différences relatives aux proportions de l'arrière du crâne peuvent être considérées comme un artefact du fait que la plupart des spécimens de S. sanjuanensis n'étaient pas complètement développés avant la découverte des "amoureux de Tambach", qui étaient des membres adultes de les espèces.
Néanmoins, plusieurs caractéristiques sont encore clairement différenciées entre les deux espèces. L' os lacrymal , devant les yeux, n'occupe que le bord antérieur de l'orbite chez S. baylorensis . Cependant, les spécimens de S. sanjuanensis ont une branche lacrymale qui s'étend sur une petite distance sous l'orbite. Chez S. sanjuanensis , une grande partie du bord arrière de l'orbite est formée par l' os postorbitaire en forme de chevron , qui est plus rectangulaire chez S. baylorensis . La forme du lacrymal et du postorbitaire de S. sanjuanensis correspond étroitement à l'état d'autres seymouriamorphes, tandis que l'état de S. baylorensis est plus unique et dérivé.
L'os maxillaire porteur de dents, qui forme le côté du museau, est également distinctement unique chez S. baylorensis . Chez S. sanjuanensis , le maxillaire était bas, avec de nombreuses dents pointues et rapprochées s'étendant sur toute sa longueur. Cette condition est similaire à celle des autres seymouriamorphes. Cependant, S. baylorensis a un museau plus haut et ses dents sont généralement beaucoup plus grandes, moins nombreuses et de taille moins homogène. Le palais est généralement similaire entre les deux espèces, bien que les ectopterygoïdes soient plus triangulaires chez S. baylorensis et rectangulaires chez S. sanjuanensis .
Paléobiologie
Mode de vie
Romer (1928) a été parmi les premiers auteurs à discuter des implications biologiques du squelette de Seymouria . Il a fait valoir que les membres robustes et le corps large soutenaient l'idée qu'il s'agissait d'un animal terrestre fort avec une démarche tentaculaire. Cependant, il a également noté que les voies ferrées du Permien soutiennent généralement l'idée que les tétrapodes terrestres de cette période n'étaient pas des dragueurs sur le ventre, mais étaient plutôt assez forts pour garder leur corps hors du sol. Comme d'autres paléontologues de l'époque, Romer supposait que Seymouria avait un mode de reproduction reptilien (ou amniote), avec des œufs pondus sur la terre ferme et protégés des éléments par une membrane amniotique .
White (1939) a élaboré sur les implications biologiques. Il a noté que la présence d'une encoche otique réduit la force de la mâchoire en diminuant la quantité de surface que les muscles de la mâchoire peuvent attacher à l'intérieur du crâne. De plus, le crâne aurait été plus fragile en raison de la présence d'une si grande incision. Dans l'ensemble, il a trouvé peu probable que Seymouria soit capable de s'attaquer à de grandes proies actives. Néanmoins, les sites d'attache musculaire du palais étaient plus développés que ceux des amphibiens contemporains. White a extrapolé que Seymouria était un généraliste principalement carnivore et omnivore, se nourrissant d'invertébrés, de petits poissons et peut-être même de matériel végétal. Il peut même avoir été cannibale selon ses calculs.
White a également attiré l'attention sur les vertèbres enflées inhabituelles, qui auraient facilité le mouvement latéral (côté à côté) mais interdiraient toute torsion (torsion) de la colonne vertébrale. Cela aurait été bénéfique, car Seymouria avait des membres bas et un corps large et lourd qui aurait autrement été vulnérable à la torsion lorsqu'il marchait. Cela peut également expliquer la présence de ce trait chez les captorhinides, les diadectomorphes et autres « cotylosaures ». Peut-être que les vertèbres enflées étaient une stratégie provisoire pour empêcher la torsion, qui serait plus tard supplantée par des muscles de la hanche puissants chez les reptiles ultérieurs. Les muscles de la hanche assez peu développés de Seymouria sont en ligne avec cette hypothèse. Néanmoins, ces vertèbres étaient inefficaces pour se défendre contre la torsion à n'importe quelle vitesse plus rapide qu'une marche rapide, donc Seymouria n'était probablement pas un animal se déplaçant rapidement.
Bien que White considérait Seymoria comme assez compétent sur terre, il a également discuté de quelques autres modes de vie. Il supposait que Seymouria était aussi un bon nageur, puisqu'il estimait (à tort) que l'animal avait une queue profonde et puissante semblable à celle des crocodiliens modernes . Cependant, il a également noté qu'il aurait été vulnérable aux prédateurs semi-aquatiques ou aquatiques, et que les fossiles de Seymouria étaient plus fréquents dans les dépôts terrestres en raison de ses préférences en matière d'habitat. Berman et al . (2000) ont soutenu cette hypothèse, car la Formation de Tambach a conservé des fossiles de Seymouria tout en manquant complètement d'animaux aquatiques. Ils ont également souligné que les os du poignet et de la cheville bien développés des "amoureux de Tambach" soutenaient les affinités terrestres. Malgré la forte musculature des membres antérieurs, Romer (1928) et White (1939) n'ont trouvé que peu de preuves d'adaptations à l' enfouissement chez Seymouria .
Dimorphisme sexuel
Certains auteurs ont plaidé en faveur du dimorphisme sexuel existant chez Seymouria , mais d'autres ne sont pas convaincus par cette hypothèse. White (1939) a soutenu que certains spécimens de Seymouria baylorensis avaient des chevrons (épines osseuses sur la face inférieure des vertèbres caudales) qui apparaissaient d'abord sur la troisième vertèbre caudale, tandis que d'autres spécimens les faisaient apparaître pour la première fois sur la sixième. Il a postulé que l'apparition ultérieure des chevrons dans certains spécimens indiquait qu'il s'agissait de mâles ayant besoin de plus d'espace pour stocker leurs organes génitaux internes. Ce type de différenciation sexuelle a été signalé chez les tortues et les crocodiliens. Sur cette base, il a également soutenu l'idée que les femelles Seymouria donnaient naissance à des œufs à gros jaune sur terre, comme les tortues et les crocodiliens. Vaughn (1966) a trouvé plus tard une corrélation entre l'acquisition de chevrons et certaines proportions du crâne chez Seymouria sanjuanensis , et a proposé qu'ils étaient aussi des exemples de dimorphisme sexuel.
Cependant, Berman, Reisz et Elberth (1987) ont critiqué les méthodologies de White (1939) et de Vaughn (1966). Ils ont fait valoir que les observations de White n'étaient probablement pas liées au sexe des animaux. Cela a été soutenu par le fait que certains des spécimens de la formation Cutler avaient des chevrons qui sont apparus pour la première fois sur leur cinquième vertèbre caudale. Bien qu'il soit possible que la taille des organes génitaux soit variable chez les mâles au point d'avoir un impact sur le squelette, l'explication la plus probable était que les différences observées par White étaient causées par une variation squelettique individuelle, une divergence évolutive ou un autre facteur sans rapport avec le dimorphisme sexuel. De même, ils ont convenu que les proportions du crâne soutenaient la proposition de Vaughn (1966) selon laquelle le dimorphisme était présent dans les fossiles de Seymouria , bien qu'ils n'étaient pas d'accord avec la façon dont il l'avait lié au sexe en utilisant un fossile qui était considéré comme « femelle » selon les critères de White. La découverte de seymouriamorphes larvaires fossilisés a montré que Seymouria avait probablement un stade larvaire aquatique, démystifiant les hypothèses antérieures selon lesquelles Seymouria pondait des œufs sur terre.
Histologie et développement
Des preuves histologiques provenant de spécimens trouvés à Richards Spurs , Oklahoma, ont fourni des informations supplémentaires sur la biologie de Seymouria . Un fémur s'est avéré avoir une structure interne caractérisée par une matrice lamellaire percée de nombreux canaux plexiformes. Les lignes de repos de croissance lente sont indistinctes et rapprochées, mais il n'y a aucune preuve que la croissance ait cessé à un moment quelconque au cours du développement osseux. Comme la plupart des lissamphibiens , la cavité médullaire est ouverte et contient une petite quantité d'os spongieux. Le développement de l'os spongieux est légèrement supérieur à celui de l' Acheloma (un amphibien terrestre), mais est beaucoup moins étendu que celui des amphibiens aquatiques tels que Rhinesuchus et Trimerorhachis . Les vertèbres de Seymouria sont de forme plus robuste que celles de Discosauriscus et ont une faible quantité de cartilage malgré une grande porosité. On suppose que Seymouria a subi une métamorphose très tôt dans sa vie, probablement en raison des stress environnementaux causés par les saisons sèches et humides fluctuantes.
Les références
Liens externes
- Une photographie du spécimen "Tambach lovers", publiée par le compte twitter de Mark MacDougall
- Une autre photographie des "Amants de Tambach", publiée par "Geology Page"
- Une photographie du bloc Cutler Formation, publiée par le compte twitter de "mskvarla36"
- Documentaire traduit de DW sur les fossiles de Tambach, dont Seymouria
{kind=link}
{kind=link}
{kind=link}