
Pilus - Pilus

Un pilus ( latin pour 'cheveux' ; pluriel : pili ) est un appendice ressemblant à un cheveu que l'on trouve à la surface de nombreuses bactéries et archées . Les termes pilus et fimbria (latin pour « frange » ; pluriel : fimbriae ) peuvent être utilisés de manière interchangeable, bien que certains chercheurs réservent le terme pilus pour l'appendice requis pour la conjugaison bactérienne . Tous les pili dans ce dernier sens sont principalement composés de protéines piline , qui sont oligomères .
Des dizaines de ces structures peuvent exister à la surface des bactéries et des archées. Certaines bactéries, virus ou bactériophages se fixent aux récepteurs des pili au début de leur cycle de reproduction .
Les pili sont antigéniques . Ils sont également fragiles et constamment remplacés, parfois par des pili de composition différente, entraînant une altération de l'antigénicité. Les réponses spécifiques de l'hôte à l'ancienne structure pili ne sont pas efficaces sur la nouvelle structure. Les gènes de recombinaison des pili codent pour les régions variables (V) et constantes (C) des pili (similaire à la diversité des immunoglobulines ). En tant que principaux déterminants antigéniques, facteurs de virulence et facteurs d'impunité à la surface cellulaire d'un certain nombre d'espèces de bactéries à Gram négatif et de certaines bactéries à Gram positif, y compris les Enterobacteriaceae , les Pseudomonadaceae et les Neisseriaceae , l'étude des pili en tant qu'organite de adhésion et comme composants du vaccin. La première étude détaillée des pili a été réalisée par Brinton et ses collègues qui ont démontré l'existence de deux phases distinctes au sein d'une souche bactérienne : pile (p+) et non pile)
Types par fonction
Quelques noms sont donnés à différents types de pili par leur fonction. La classification ne chevauche pas toujours les types structurels ou évolutifs, car une évolution convergente se produit.
Pili conjugatif
Les pili conjugatifs permettent le transfert d' ADN entre bactéries, dans le processus de conjugaison bactérienne . Ils sont parfois appelés « sex pili », par analogie à la reproduction sexuée , car ils permettent l'échange de gènes via la formation de « couples d'accouplement ». Peut-être le plus bien étudié est le F-pili d' Escherichia coli , codée par le facteur sexuel F .
Un pilus sexuel a généralement un diamètre de 6 à 7 nm . Au cours de la conjugaison, un pilus émergeant de la bactérie donneuse piège la bactérie receveuse, la rapproche et déclenche finalement la formation d'un pont d'accouplement , qui établit un contact direct et la formation d'un pore contrôlé qui permet le transfert d'ADN du donneur à le destinataire. Typiquement, l'ADN transféré se compose des gènes nécessaires à la fabrication et au transfert de pili (souvent codés sur un plasmide ), et il s'agit donc d'une sorte d' ADN égoïste ; cependant, d'autres morceaux d'ADN sont souvent co-transférés, ce qui peut entraîner la dissémination de traits génétiques dans une population bactérienne, comme la résistance aux antibiotiques . Toutes les bactéries ne peuvent pas produire de pili conjugatif, mais la conjugaison peut se produire entre des bactéries d'espèces différentes.
Fimbriae
Fimbria est un terme utilisé pour un pilus court qui est utilisé pour attacher la bactérie à une surface, parfois aussi appelé « pilus d'attachement ». Les fimbriae sont nécessaires à la formation du biofilm, car elles attachent les bactéries aux surfaces hôtes pour la colonisation pendant l'infection. Les fimbriae sont soit situés aux pôles d'une cellule, soit uniformément répartis sur toute sa surface. Les bactéries mutantes dépourvues de fimbriae ne peuvent pas adhérer à leurs surfaces cibles habituelles et ne peuvent donc pas provoquer de maladies .
Ce terme a également été utilisé dans un sens laxiste pour désigner tous les pili, par ceux qui utilisent « pilus » pour désigner spécifiquement les pili sexuels.
Types par système d'assemblage ou structure
Transfert
La famille Tra (transfert) comprend tous les pili sexuels connus (en 2010). Ils sont liés au système de sécrétion de type IV (T4SS). Ils peuvent être classés dans le type F-like (après le F-pilus) et le type P-like. Comme leurs homologues de sécrétion, le pilus injecte du matériel, de l'ADN dans ce cas, dans une autre cellule.
Pili de type IV

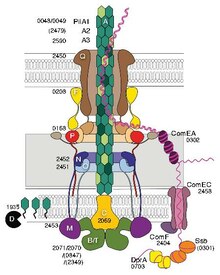
Certains pili, appelés pili de type IV (T4P), génèrent des forces mobiles . Les extrémités externes des pili adhèrent à un substrat solide, soit à la surface à laquelle la bactérie est attachée, soit à d'autres bactéries. Ensuite, lorsque les pili se contractent, ils tirent la bactérie vers l'avant comme un grappin. Le mouvement produit par les pili de type IV est généralement saccadé, c'est pourquoi on l'appelle motilité convulsive , par opposition à d'autres formes de motilité bactérienne telles que celle produite par les flagelles . Cependant, certaines bactéries, par exemple Myxococcus xanthus , présentent une motilité glissante . Les pili bactériens de type IV ont une structure similaire aux flagellines constitutives de l' archaella (flagelles archées ), et les deux sont liés au système de sécrétion de type II (T2SS); ils sont unifiés par le groupe des systèmes de filaments de type IV . Outre les archées, de nombreuses archées produisent des pili adhésifs de type 4, qui permettent aux cellules archéennes d'adhérer à différents substrats. Les parties alpha-hélicoïdales N-terminales des pilines et archaellines archées de type 4 sont homologues aux régions correspondantes de la T4P bactérienne; cependant, les domaines riches en brins bêta C-terminaux semblent être sans rapport dans les pilines bactériennes et archéennes.
La transformation génétique est le processus par lequel une cellule bactérienne réceptrice prélève l'ADN d'une cellule voisine et intègre cet ADN dans son génome par recombinaison homologue . Chez Neisseria meningitidis (également appelé méningocoque), la transformation de l'ADN nécessite la présence de courtes séquences d'absorption d'ADN (DUS) qui sont 9 à 10 monomères résidant dans les régions codantes de l'ADN du donneur. La reconnaissance spécifique des DHS est médiée par une piline de type IV . Les pili du méningocoque de type IV se lient à l'ADN par le biais de la ComP mineure de piline via une bande électropositive qui devrait être exposée à la surface du filament. ComP affiche une préférence de reliure exquise pour les DHS sélectifs. La distribution des DUS au sein du génome de N. meningitides favorise certains gènes, ce qui suggère qu'il existe un biais pour les gènes impliqués dans la maintenance et la réparation génomiques.
Cette famille a été identifiée à l'origine comme « fimbriae de type IV » par leur apparition au microscope. Cette classification a survécu car elle correspond à un clade.
Virulence
Les pili sont responsables de la virulence des souches pathogènes de nombreuses bactéries, dont E. coli , Vibrio cholerae et de nombreuses souches de Streptococcus . En effet, la présence de pili améliore considérablement la capacité des bactéries à se lier aux tissus corporels, ce qui augmente ensuite les taux de réplication et la capacité d'interagir avec l'organisme hôte. Si une espèce de bactérie a plusieurs souches mais que seules certaines sont pathogènes, il est probable que les souches pathogènes auront des pili alors que les souches non pathogènes n'en auront pas.
Le développement de pili d'attachement peut alors entraîner le développement d'autres traits de virulence. Les souches non pathogènes de V. cholerae ont d' abord développé des pili, leur permettant de se lier aux tissus humains et de former des microcolonies . Ces pili ont ensuite servi de sites de liaison pour le bactériophage lysogène qui transporte la toxine causant la maladie . Le gène de cette toxine, une fois incorporé dans le génome de la bactérie, est exprimé lors de l'expression du gène codant pour le pilus (d'où le nom de « toxine mediated pilus »).
Voir également
Les références
Liens externes
- Sex+Pilus à la National Library of Medicine Medical Subject Headings (MeSH) des États-Unis
- Bacterial+Pilus à la National Library of Medicine Medical Subject Headings (MeSH) des États-Unis