
Plasmodesme - Plasmodesma

Les plasmodesmes (singulier : plasmodesma ) sont des canaux microscopiques qui traversent les parois cellulaires des cellules végétales et de certaines cellules d' algues , permettant le transport et la communication entre elles. Les plasmodesmes ont évolué indépendamment dans plusieurs lignées, et les espèces qui possèdent ces structures comprennent des membres des Charophyceae , des Charales , des Coleochaetales et des Phaeophyceae (qui sont toutes des algues), ainsi que tous les embryophytes , mieux connus sous le nom de plantes terrestres. Contrairement aux cellules animales , presque toutes les cellules végétales sont entourées d'un polysaccharide paroi cellulaire . Les cellules végétales voisines sont donc séparées par une paire de parois cellulaires et la lamelle intermédiaire intermédiaire , formant un domaine extracellulaire connu sous le nom d' apoplaste . Bien que les parois cellulaires soient perméables aux petites protéines solubles et autres solutés , les plasmodesmes permettent le transport direct, régulé et symplastique de substances entre les cellules. Il existe deux formes de plasmodesmes : les plasmodesmes primaires, qui se forment lors de la division cellulaire, et les plasmodesmes secondaires, qui peuvent se former entre les cellules matures.
Des structures similaires, appelées jonctions communicantes et nanotubes membranaires , interconnectent les cellules animales et les stromos se forment entre les plastes des cellules végétales.
Formation
Les plasmodesmes primaires se forment lorsque des fractions du réticulum endoplasmique sont piégées à travers la lamelle moyenne alors qu'une nouvelle paroi cellulaire est synthétisée entre deux cellules végétales nouvellement divisées. Ceux-ci finissent par devenir les connexions cytoplasmiques entre les cellules. Sur le site de formation, la paroi ne s'épaissit pas davantage et des dépressions ou des zones minces appelées fosses se forment dans les parois. Les fosses s'apparient normalement entre les cellules adjacentes. Les plasmodesmes peuvent également être insérés dans les parois cellulaires existantes entre les cellules qui ne se divisent pas (plasmodesmes secondaires).
Plasmodesmes primaires
La formation de plasmodesmes primaires se produit pendant la partie du processus de division cellulaire où le réticulum endoplasmique et la nouvelle plaque sont fusionnés, ce processus entraîne la formation d'un pore cytoplasmique (ou manchon cytoplasmique). Le desmotubule, également connu sous le nom de RE apprimé, se forme à côté du RE cortical. Le RE apprimé et le RE cortical sont tous deux étroitement emballés, ne laissant ainsi aucune place à l'espace luminal. Il est proposé que le RE apprimé agisse comme une voie de transport membranaire dans les plasmodesmes. Lorsque les filaments du RE cortical sont enchevêtrés dans la formation d'une nouvelle plaque cellulaire, la formation de plasmodesmes se produit chez les plantes terrestres. On suppose que le RE apprimé se forme en raison d'une combinaison de la pression d'une paroi cellulaire en croissance et de l'interaction des protéines ER et PM. Les plasmodesmes primaires sont souvent présents dans les zones où les parois cellulaires semblent être plus minces. Cela est dû au fait qu'à mesure qu'une paroi cellulaire se dilate, l'abondance des plasmodesmes primaires diminue. Afin d'augmenter davantage la densité plasmodesmale pendant la croissance de la paroi cellulaire, des plasmodesmes secondaires sont produits. Le processus de formation des plasmodesmes secondaires n'est pas encore entièrement compris, mais diverses enzymes de dégradation et protéines ER sont censées stimuler le processus.
Structure
Membrane plasmique plasmodesmateuse
Une cellule végétale typique peut avoir entre 10 3 et 10 5 plasmodesmes la reliant à des cellules adjacentes, ce qui équivaut à entre 1 et 10 par µm 2 . Les plasmodesmes mesurent environ 50 à 60 nm de diamètre au milieu et sont constitués de trois couches principales, la membrane plasmique , le manchon cytoplasmique et le desmotubule . Ils peuvent traverser des parois cellulaires pouvant atteindre 90 nm d'épaisseur.
La partie de la membrane plasmique du plasmodesme est une extension continue de la membrane cellulaire ou du plasmalemme et a une structure bicouche phospholipidique similaire .
Le manchon cytoplasmique est un espace rempli de liquide délimité par le plasmalemme et est une extension continue du cytosol . Le trafic de molécules et d' ions à travers les plasmodesmes se produit à travers cet espace. Les molécules plus petites (par exemple les sucres et les acides aminés ) et les ions peuvent facilement traverser les plasmodesmes par diffusion sans avoir besoin d'énergie chimique supplémentaire. Les molécules plus grosses, y compris les protéines (par exemple la protéine fluorescente verte ) et l'ARN, peuvent également traverser le manchon cytoplasmique de manière diffuse. Le transport plasmodesmatal de certaines molécules plus grosses est facilité par des mécanismes actuellement inconnus. Un mécanisme de régulation de la perméabilité des plasmodesmes est l'accumulation du polysaccharide callose autour de la région du cou pour former une collerette, réduisant ainsi le diamètre du pore disponible pour le transport des substances. Grâce à la dilatation, la synchronisation active ou le remodelage structurel, la perméabilité des plasmodesmes est augmentée. Cette augmentation de la perméabilité des pores des plasmodesmes permet de transporter des molécules plus grosses, ou macromolécules , telles que des molécules de signalisation, des facteurs de transcription et des complexes ARN-protéine vers divers compartiments cellulaires.
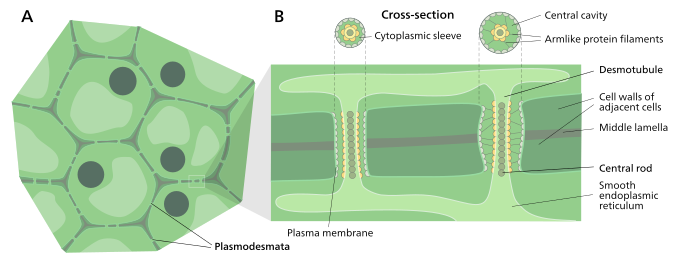
Structure d'un plasmodesme et sa localisation dans les cellules végétales

Les plasmodesmes permettent aux molécules de voyager entre les cellules végétales par la voie symplastique


Desmotubule
Le desmotubule est un tube de apprimées (aplatie) réticulum endoplasmique qui passe entre deux cellules adjacentes. Certaines molécules sont connues pour être transportées par ce canal, mais on ne pense pas qu'il s'agisse de la principale voie de transport plasmodesmatique.
Autour du desmotubule et de la membrane plasmique, des zones d'un matériau dense aux électrons ont été observées, souvent reliées entre elles par des structures en forme de rayons qui semblent diviser le plasmodesma en canaux plus petits. Ces structures peuvent être composées de myosine et d' actine , qui font partie du cytosquelette de la cellule . Si tel est le cas, ces protéines pourraient être utilisées dans le transport sélectif de grosses molécules entre les deux cellules.
Transport

Plasmodesmes On a montré que le transport des protéines (y compris les facteurs de transcription ), ARN interférant court , ARN messenger , viroïdes , virus et génomes de cellule à cellule. Un exemple de protéines de mouvement viral est le virus de la mosaïque du tabac MP-30. On pense que le MP-30 se lie au propre génome du virus et le transporte des cellules infectées vers les cellules non infectées via les plasmodesmes. La protéine du Locus T de floraison se déplace des feuilles vers le méristème apical de la pousse en passant par les plasmodesmes pour initier la floraison .
Les plasmodesmes sont également utilisés par les cellules du phloème et le transport symplastique est utilisé pour réguler les cellules du tube criblé par les cellules compagnes .
La taille des molécules qui peuvent traverser les plasmodesmes est déterminée par la limite d'exclusion de taille. Cette limite est très variable et est sujette à modification active. Par exemple, le MP-30 est capable d'augmenter la limite d'exclusion de taille de 700 Daltons à 9400 Daltons, facilitant ainsi son déplacement dans une usine. En outre, il a été démontré que l'augmentation des concentrations de calcium dans le cytoplasme, soit par injection, soit par induction par le froid, restreint l'ouverture des plasmodesmes environnants et limite le transport.
Il existe plusieurs modèles de transport actif possible à travers les plasmodesmes. Il a été suggéré qu'un tel transport est médié par des interactions avec des protéines localisées sur le desmotubule, et/ou par des chaperons dépliant partiellement des protéines , leur permettant de s'adapter à travers le passage étroit. Un mécanisme similaire peut être impliqué dans le transport des acides nucléiques viraux à travers les plasmodesmes.
Un certain nombre de modèles mathématiques ont été suggérés pour estimer le transport à travers les plasmodesmes. Ces modèles ont principalement traité le transport comme un problème de diffusion avec quelques obstacles supplémentaires.
Composants cytosquelettiques des Plasmodesmes
Les plasmodesmes relient presque toutes les cellules d'une plante, ce qui peut provoquer des effets négatifs tels que la propagation de virus. Pour comprendre cela, nous devons d'abord examiner les composants du cytosquelette, tels que les microfilaments d'actine, les microtubules et les protéines de la myosine, et comment ils sont liés au transport de cellule à cellule. Les microfilaments d'actine sont liés au transport des protéines de mouvement viral vers les plasmodesmes qui permettent le transport de cellule à cellule à travers les plasmodesmes. Le marquage fluorescent pour la co-expression dans les feuilles de tabac a montré que les filaments d'actine sont responsables du transport des protéines de mouvement viral vers les plasmodesmes. Lorsque la polymérisation de l'actine a été bloquée, elle a provoqué une diminution du ciblage des plasmodesmes des protéines de mouvement dans le tabac et a permis aux composants de 10 kDa (plutôt que de 126 kDa) de se déplacer entre les cellules du mésophylle du tabac . Cela a également eu un impact sur le mouvement de cellule à cellule des molécules dans la plante de tabac.
Virus
Les virus décomposent les filaments d'actine dans le canal des plasmodesmes afin de se déplacer à l'intérieur de la plante. Par exemple, lorsque le virus de la mosaïque du concombre (CMV) pénètre dans les plantes, il est capable de traverser presque toutes les cellules grâce à l'utilisation de protéines de mouvement viral pour se transporter à travers les plasmodesmes. Lorsque les feuilles de tabac sont traitées avec un médicament qui stabilise les filaments d'actine, la phalloïdine, les protéines de mouvement du virus de la mosaïque du concombre sont incapables d'augmenter la limite d'exclusion de taille des plasmodesmes (SEL).
Myosine
Des quantités élevées de protéines de myosine se trouvent sur les sites des plasmodesmes. Ces protéines sont impliquées dans la direction des cargaisons virales vers les plasmodesmes. Lorsque des formes mutantes de myosine ont été testées dans des plants de tabac, le ciblage des protéines virales vers les plasmodesmes a été affecté négativement. La liaison permanente de la myosine à l'actine, induite par un médicament, a entraîné une diminution du mouvement de cellule à cellule. Les virus sont également capables de se lier sélectivement aux protéines de la myosine.
Microtubules
Les microtubules ont un rôle important dans le transport de cellule à cellule de l'ARN viral. Les virus utilisent de nombreuses méthodes différentes pour se transporter d'une cellule à l'autre et l'une de ces méthodes associe le domaine N-terminal de son ARN pour se localiser dans les plasmodesmes à travers les microtubules. Dans les plants de tabac injectés avec des virus de la mosaïque du tabac qui ont été maintenus à des températures élevées, il y avait une forte corrélation entre les protéines de mouvement du TMV marquées par la GFP et les microtubules. Cela a conduit à une augmentation de la propagation de l'ARN viral à travers le tabac.
Plasmodesmes et callose
La régulation et la structure des plasmodesmes sont régulées par un polymère bêta 1,3-glucane appelé callose. La callose se trouve dans les plaques cellulaires pendant le processus de cytokinèse, mais à mesure que ce processus arrive à son terme, les niveaux de callose diminuent. Les seules parties riches en callose de la cellule comprennent les sections de la paroi cellulaire où sont présents les plasmodesmes. Afin de réguler ce qui est transporté à travers les plasmodesmes, la callose doit être présente. La callose fournit le mécanisme par lequel la perméabilité des plasmodesmes est régulée. Afin de contrôler ce qui est transporté entre les différents tissus, les plasmodesmes subissent plusieurs changements de conformation spécialisés.
L'activité des plasmodesmes est liée aux processus physiologiques et développementaux des plantes. Il existe une voie de signalisation hormonale qui relaie les signaux cellulaires primaires via les plasmodesmes. Il existe également des modèles d'indices environnementaux, physiologiques et développementaux qui montrent une relation avec la fonction des plasmodesmes. Un mécanisme important des plasmodesmes est la capacité d'ouvrir ses canaux. Les niveaux de callose se sont avérés être une méthode pour changer la taille de l'ouverture des plasmodesmes. Des dépôts de callose se trouvent au col des plasmodesmes dans les nouvelles parois cellulaires qui se sont formées. Le niveau de dépôts au niveau des plasmodesmes peut fluctuer, ce qui montre qu'il existe des signaux qui déclenchent une accumulation de callose au niveau des plasmodesmes et provoquent l'ouverture ou l'ouverture des plasmodesmes. Les activités enzymatiques de la bêta 1,3-glucane synthase et des hydrolases sont impliquées dans les changements du niveau de cellulose des plasmodesmes. Certains signaux extracellulaires modifient la transcription des activités de cette synthase et hydrolase. Arabidopsis thaliana possède des gènes de callose synthase qui codent pour une sous-unité catalytique du B-1,3-glucane. Les mutants à gain de fonction dans ce pool génétique montrent un dépôt accru de callose au niveau des plasmodesmes et une diminution du trafic macromoléculaire ainsi qu'un système racinaire défectueux au cours du développement.