
Vol d'insectes - Insect flight

Les insectes sont le seul groupe d' invertébrés qui ont développé des ailes et un vol . Les insectes ont volé pour la première fois au Carbonifère , il y a 350 à 400 millions d'années, faisant d'eux les premiers animaux à évoluer en vol. Les ailes peuvent avoir évolué à partir d'appendices sur les côtés des membres existants, qui avaient déjà des nerfs, des articulations et des muscles utilisés à d'autres fins. Ceux-ci peuvent avoir été initialement utilisés pour naviguer sur l'eau, ou pour ralentir le taux de descente en planant.
Deux groupes d'insectes, les libellules et les éphémères , ont des muscles de vol attachés directement aux ailes. Chez d'autres insectes ailés, les muscles du vol s'attachent au thorax, ce qui le fait osciller afin d'inciter les ailes à battre. Parmi ces insectes, certains ( mouches et certains coléoptères ) atteignent des fréquences de battement d'ailes très élevées grâce à l'évolution d'un système nerveux "asynchrone", dans lequel le thorax oscille plus rapidement que le rythme des impulsions nerveuses.
Certains très petits insectes n'utilisent pas l' aérodynamique en régime permanent , mais le mécanisme de clap et de projection de Weis-Fogh , générant de grandes forces de portance au détriment de l'usure des ailes. De nombreux insectes peuvent planer, maintenir leur hauteur et contrôler leur position. Certains insectes tels que les mites ont les ailes antérieures couplées aux ailes postérieures afin qu'elles puissent travailler à l'unisson.
Mécanismes
Vol direct
Contrairement à d'autres insectes, les muscles des ailes des éphéméroptères (éphémères) et des odonates (libellules et demoiselles) s'insèrent directement à la base des ailes, qui sont articulées de sorte qu'un petit mouvement vers le bas de la base de l'aile soulève l'aile elle-même vers le haut, un peu comme ramer à travers l'air. Les libellules et les demoiselles ont des ailes antérieures et postérieures de forme et de taille similaires. Chacun fonctionne indépendamment, ce qui donne un degré de contrôle fin et de mobilité en termes de brusquerie avec laquelle ils peuvent changer de direction et de vitesse, que l'on ne voit pas chez d'autres insectes volants. Les odonates sont tous des prédateurs aériens et ils ont toujours chassé d'autres insectes aériens.

Vol direct : muscles attachés aux ailes. Gros insectes seulement


Vol indirect
À part les deux ordres avec des muscles de vol directs, tous les autres insectes ailés vivants volent en utilisant un mécanisme différent, impliquant des muscles de vol indirects. Ce mécanisme a évolué une fois et est la caractéristique déterminante ( synapomorphie ) pour l' infraclasse Neoptera ; cela correspond, probablement pas par hasard, à l'apparition d'un mécanisme de pliage des ailes, qui permet aux insectes néoptères de replier les ailes sur l'abdomen au repos (bien que cette capacité ait été perdue secondairement dans certains groupes, comme chez les papillons ) .
Ce que tous les Néoptères partagent, cependant, c'est la façon dont fonctionnent les muscles du thorax : ces muscles, plutôt que de s'attacher aux ailes, s'attachent au thorax et le déforment ; comme les ailes sont des extensions de l' exosquelette thoracique , les déformations du thorax font également bouger les ailes. Un ensemble de muscles longitudinaux le long du dos comprime le thorax d'avant en arrière, provoquant la courbure de la surface dorsale du thorax ( notum ) vers le haut, faisant basculer les ailes vers le bas. Un autre ensemble de muscles du tergum au sternum tire à nouveau le notum vers le bas, provoquant le basculement des ailes vers le haut.
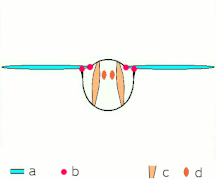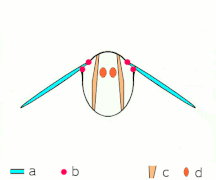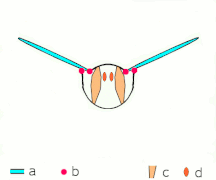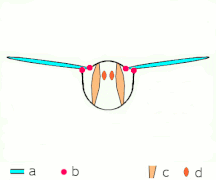
Vol indirect : les muscles font osciller le thorax chez la plupart des insectes


Les insectes qui battent des ailes moins de cent fois par seconde utilisent des muscles synchrones. Le muscle synchrone est un type de muscle qui se contracte une fois pour chaque influx nerveux. Cela produit généralement moins de puissance et est moins efficace que le muscle asynchrone, ce qui explique l'évolution indépendante des muscles de vol asynchrones dans plusieurs clades d'insectes distincts.
Les insectes qui battent des ailes plus rapidement, comme le bourdon , utilisent le muscle asynchrone ; c'est un type de muscle qui se contracte plus d'une fois par impulsion nerveuse. Pour ce faire, le muscle est stimulé pour se contracter à nouveau par un relâchement de la tension dans le muscle, ce qui peut se produire plus rapidement que par une simple stimulation nerveuse seule. Cela permet à la fréquence des battements d'ailes de dépasser la vitesse à laquelle le système nerveux peut envoyer des impulsions. Le muscle asynchrone est l'un des derniers raffinements apparus chez certains des néoptères supérieurs ( coléoptères , diptères et hyménoptères ). L'effet global est que de nombreux Néoptères supérieurs peuvent battre des ailes beaucoup plus rapidement que les insectes avec des muscles de vol directs.
Aérodynamique
Il existe deux modèles aérodynamiques de base du vol des insectes : la création d'un vortex de bord d'attaque et l'utilisation du clap and fling.
Vortex de bord d'attaque
La plupart des insectes utilisent une méthode qui crée un vortex de bord d'attaque en spirale . Ces ailes battantes se déplacent en deux demi-courses de base. La course descendante démarre et recule et est plongée vers le bas et vers l'avant. Puis l'aile est rapidement retournée ( supination ) de sorte que le bord d'attaque soit pointé vers l'arrière. La course ascendante pousse alors l'aile vers le haut et vers l'arrière. Ensuite, l'aile est à nouveau retournée ( pronation ) et une autre descente peut se produire. La gamme de fréquences chez les insectes avec des muscles de vol synchrones est généralement de 5 à 200 hertz (Hz). Chez ceux qui ont des muscles de vol asynchrones, la fréquence des battements d'ailes peut dépasser 1000 Hz. Lorsque l'insecte est en vol stationnaire, les deux coups prennent le même temps. Une course descendante plus lente, cependant, fournit une poussée .
L'identification des principales forces est essentielle pour comprendre le vol des insectes. Les premières tentatives pour comprendre le battement d'ailes supposaient un état quasi-stationnaire. Cela signifie que le flux d'air au-dessus de l'aile à un moment donné était supposé être le même que le flux d'air au-dessus d'une aile stable et non battante au même angle d'attaque. En divisant l'aile battante en un grand nombre de positions immobiles puis en analysant chaque position, il serait possible de créer une chronologie des forces instantanées sur l'aile à chaque instant. La portance calculée s'est avérée trop petite d'un facteur trois, les chercheurs ont donc réalisé qu'il devait y avoir des phénomènes instables fournissant des forces aérodynamiques. Il y avait plusieurs modèles analytiques en développement tentant d'approcher le flux près d'une aile battante. Certains chercheurs ont prédit des pics de force lors de la supination. Avec un modèle à l'échelle dynamique d'une mouche des fruits , ces forces prédites ont été confirmées plus tard. D'autres ont soutenu que les pics de force pendant la supination et la pronation sont causés par un effet de rotation inconnu qui est fondamentalement différent des phénomènes de translation. Il y a un certain désaccord avec cet argument. Grâce à la dynamique des fluides computationnelle , certains chercheurs soutiennent qu'il n'y a pas d'effet rotationnel. Ils prétendent que les forces élevées sont causées par une interaction avec le sillage du coup précédent.
Semblable à l'effet de rotation mentionné ci-dessus, les phénomènes associés au battement d'ailes ne sont pas complètement compris ou acceptés. Parce que chaque modèle est une approximation, différents modèles laissent de côté des effets qui sont présumés négligeables. Par exemple, l' effet Wagner , tel que proposé par Herbert A. Wagner en 1925, indique que la circulation augmente lentement jusqu'à son état d'équilibre en raison de la viscosité lorsqu'une aile inclinée est accélérée depuis le repos. Ce phénomène expliquerait une valeur de lift inférieure à ce qui est prédit. En règle générale, il s'agissait de trouver des sources pour l'augmentation supplémentaire. Il a été avancé que cet effet est négligeable pour un écoulement avec un nombre de Reynolds typique du vol des insectes. Le nombre de Reynolds est une mesure de la turbulence ; l'écoulement est laminaire (lisse) lorsque le nombre de Reynolds est faible, et turbulent lorsqu'il est élevé. L'effet Wagner a été ignoré, consciemment, dans au moins un modèle. L'un des phénomènes les plus importants qui se produisent pendant le vol des insectes est la succion du bord d'attaque. Cette force est importante pour le calcul de l'efficacité. Le concept d'aspiration du bord d'attaque a été proposé pour la première fois par DG Ellis et JL Stollery en 1988 pour décrire la portance vortex sur les ailes delta à arêtes vives . Aux angles d'attaque élevés, le flux se sépare sur le bord d'attaque, mais se rattache avant d'atteindre le bord de fuite. A l'intérieur de cette bulle d'écoulement séparé se trouve un vortex. Parce que l'angle d'attaque est si élevé, beaucoup d'élan est transféré vers le bas dans le flux. Ces deux caractéristiques créent une force de portance importante ainsi qu'une traînée supplémentaire. La caractéristique importante, cependant, est l'ascenseur. Parce que le flux s'est séparé, mais qu'il fournit toujours de grandes quantités de portance, ce phénomène est appelé retard de décrochage , remarqué pour la première fois sur les hélices d'avions par H. Himmelskamp en 1945. Cet effet a été observé dans le vol d'insectes battants et il s'est avéré capable de fournissant suffisamment d'ascenseur pour tenir compte de la déficience dans les modèles quasi-stationnaire. Cet effet est utilisé par les canoéistes dans un coup de godille .
Tous les effets sur une aile battante peuvent être réduits à trois sources majeures de phénomènes aérodynamiques : le tourbillon de bord d'attaque, les forces aérodynamiques en régime permanent sur l'aile et le contact de l'aile avec son sillage des coups précédents. La taille des insectes volants varie d'environ 20 microgrammes à environ 3 grammes. À mesure que la masse corporelle de l'insecte augmente, la surface des ailes augmente et la fréquence des battements d'ailes diminue. Pour les insectes plus gros, le nombre de Reynolds (Re) peut atteindre 10 000, là où le flux commence à devenir turbulent. Pour les insectes plus petits, il peut être aussi bas que 10. Cela signifie que les effets visqueux sont beaucoup plus importants pour les insectes plus petits.


Une autre caractéristique intéressante du vol des insectes est l'inclinaison du corps. À mesure que la vitesse de vol augmente, le corps de l'insecte a tendance à s'incliner vers le bas et à devenir plus horizontal. Cela réduit la zone frontale et donc la traînée du corps. Étant donné que la traînée augmente également à mesure que la vitesse vers l'avant augmente, l'insecte rend son vol plus efficace à mesure que cette efficacité devient plus nécessaire. De plus, en modifiant l'angle d'attaque géométrique lors de la descente, l'insecte est capable de maintenir son vol avec une efficacité optimale à travers autant de manœuvres que possible. Le développement de la poussée générale est relativement faible par rapport aux forces de portance. Les forces de levage peuvent être plus de trois fois le poids de l'insecte, tandis que la poussée, même aux vitesses les plus élevées, peut être aussi faible que 20 % du poids. Cette force est développée principalement par le mouvement ascendant moins puissant du mouvement de battement.
Frappez et jetez


Clap and fling, ou le mécanisme de Weis-Fogh, découvert par le zoologiste danois Torkel Weis-Fogh , est une méthode de génération de portance utilisée pendant le vol de petits insectes. Lorsque la taille des insectes devient inférieure à 1 mm, les forces visqueuses deviennent dominantes et l'efficacité de la génération de portance à partir d'un profil aérodynamique diminue considérablement. A partir de la position clap, les deux ailes s'écartent et tournent autour du bord de fuite. Les ailes se séparent alors et balayent horizontalement jusqu'à la fin de la descente. Ensuite, les ailes sont en pronation et utilisent le bord d'attaque pendant un mouvement d'aviron ascendant. Lorsque le mouvement du clap commence, les bords d'attaque se rencontrent et tournent ensemble jusqu'à ce que l'espace disparaisse. Initialement, on pensait que les ailes se touchaient, mais plusieurs incidents indiquent un écart entre les ailes et suggèrent qu'il offre un avantage aérodynamique.
La génération de portance à partir du mécanisme de claquement et de projection se produit au cours de plusieurs processus tout au long du mouvement. Premièrement, le mécanisme repose sur une interaction aile-aile, car un seul mouvement d'aile ne produit pas une portance suffisante. Lorsque les ailes tournent autour du bord de fuite dans le mouvement de projection, l'air s'engouffre dans l'espace créé et génère un puissant vortex de bord d'attaque, et un second se développe au bout des ailes. Un troisième vortex, plus faible, se développe sur le bord de fuite. La force des tourbillons en développement dépend, en partie, de l'écart initial de la séparation entre les ailes au début du mouvement de renversement. Avec un espace réduit entre les ailes indiquant une plus grande génération de portance, au prix de forces de traînée plus importantes. La mise en œuvre d'un mouvement de soulèvement pendant le vol, des ailes flexibles et un mécanisme de décrochage retardé se sont avérés renforcer la stabilité et l'attachement du vortex. Enfin, pour compenser la production globale de portance inférieure pendant le vol à faible nombre de Reynolds (avec flux laminaire ), les insectes minuscules ont souvent une fréquence de course plus élevée pour générer des vitesses de bout d'aile comparables à celles des insectes plus gros.
Les forces de traînée attendues les plus importantes se produisent pendant le mouvement de projection dorsale, car les ailes doivent se séparer et tourner. L'atténuation des grandes forces de traînée se produit par plusieurs mécanismes. On a constaté que les ailes flexibles réduisaient la traînée dans le mouvement de projection jusqu'à 50 % et réduisaient encore la traînée globale sur toute la course de l'aile par rapport aux ailes rigides. Les poils sur les bords des ailes, comme on le voit chez Encarsia formosa , provoquent une porosité dans l'écoulement qui augmente et réduit les forces de traînée, au prix d'une génération de portance plus faible. De plus, la séparation entre les ailes avant le vol joue un rôle important dans l'effet global de la traînée. Au fur et à mesure que la distance entre les ailes augmente, la traînée globale diminue.
Le mécanisme clap and fling est également utilisé par le mollusque marin Limacina helicina , un papillon marin. Certains insectes, comme la mineuse des légumes Liriomyza sativae (une mouche), exploitent un clap et un jet partiels, utilisant le mécanisme uniquement sur la partie externe de l'aile pour augmenter la portance d'environ 7 % en vol stationnaire.
- Mécanisme de vol Clap and Fling après Sane 2003

Clap 1 : les ailes se referment sur le dos

Clap 2: les bords d'attaque se touchent, l'aile tourne autour du bord d'attaque , des tourbillons se forment

Clap 3 : les bords de fuite se ferment, les tourbillons disparaissent, les ailes se ferment donnant de la poussée



- Flèches noires (courbes) : flux ; Flèches bleues : vitesse induite ; Flèches oranges : force nette sur l'aile
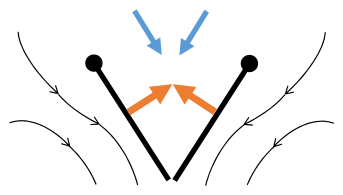
Fling 1: les ailes tournent autour du bord de fuite pour se séparer

Fling 2: le bord d'attaque s'éloigne, l'air s'engouffre, augmentant la portance
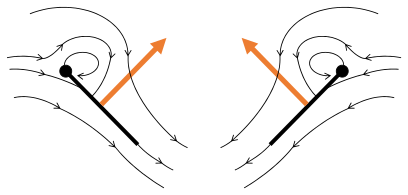
Fling 3 : de nouvelles formes de vortex au bord d'attaque, les vortex de bord de fuite s'annulent, aidant peut-être le flux à croître plus rapidement ( Weis-Fogh 1973)



Équations directrices
Une aile se déplaçant dans des fluides subit une force fluide , qui suit les conventions trouvées en aérodynamique. La composante de force normale à la direction de l'écoulement par rapport à l'aile est appelée portance ( L ), et la composante de force dans la direction opposée de l'écoulement est la traînée ( D ). Aux nombres de Reynolds considérés ici, une unité de force appropriée est 1/2(ρU 2 S), où est la densité du fluide, S la surface de l'aile et U la vitesse de l'aile. Les forces sans dimension sont appelées coefficients de portance ( C L ) et de traînée ( C D ), c'est-à-dire :

C L et C D ne sont des constantes que si le débit est constant. Une classe spéciale d'objets tels que les profils aérodynamiques peut atteindre un état stable lorsqu'elle traverse le fluide à un petit angle d'attaque. Dans ce cas, l' écoulement non visqueux autour d'un profil aérodynamique peut être approximé par un écoulement potentiel satisfaisant la condition aux limites de non-pénétration. Le théorème de Kutta-Joukowski d'un profil aérodynamique 2D suppose en outre que l'écoulement quitte le bord de fuite aigu en douceur, ce qui détermine la circulation totale autour d'un profil aérodynamique. La levée correspondante est donnée par le principe de Bernoulli ( théorème de Blasius ):

Les écoulements autour des oiseaux et des insectes peuvent être considérés comme incompressibles : le nombre de Mach , ou vitesse relative à la vitesse du son dans l'air, est typiquement de 1/300 et la fréquence des ailes est d'environ 10-103 Hz. En utilisant l'équation gouvernante comme équation de Navier-Stokes soumise à la condition aux limites sans glissement, l'équation est :

Où u (x, t) est le champ d'écoulement, p la pression, ρ la densité du fluide, ν la viscosité cinématique, u bd la vitesse à la frontière, et u est la vitesse du solide. En choisissant une échelle de longueur, L, et une échelle de vitesse, U, l'équation peut être exprimée sous une forme non dimensionnelle contenant le nombre de Reynolds, R e =uL/ν . Il y a deux différences évidentes entre une aile d'insecte et une voilure : Une aile d'insecte est beaucoup plus petite et elle bat. En utilisant une libellule comme exemple, sa corde (c) est d'environ 1 cm (0,39 in), sa longueur d'aile (l) d'environ 4 cm (1,6 in) et sa fréquence d'aile (f) d'environ 40 Hz. La vitesse de pointe (u) est d'environ 1 m/s (3,3 ft/s), et le nombre de Reynolds correspondant d'environ 103. À l'extrémité la plus petite, une guêpe Chalcid a une longueur d'aile d'environ 0,5-0,7 mm (0,020-0,028 in ) et bat son aile à environ 400 Hz. Son nombre de Reynolds est d'environ 25. La plage du nombre de Reynolds dans le vol des insectes est d'environ 10 à 10 4 , ce qui se situe entre les deux limites qui conviennent aux théories : les écoulements constants non visqueux autour d'un profil aérodynamique et l'écoulement de Stokes subi par une bactérie nageante. Pour cette raison, cette plage intermédiaire n'est pas bien comprise. D'un autre côté, c'est peut-être le régime le plus omniprésent parmi les choses que nous voyons. Les feuilles et les graines qui tombent, les poissons et les oiseaux rencontrent tous des flux instables similaires à ceux observés autour d'un insecte. Le nombre de Reynolds dans le sens de la corde peut être décrit par :

et


Où est la longueur de corde moyenne, est la vitesse du bout de l'aile, est l'amplitude de course, est la fréquence de battement, est le rayon de giration, est la surface de l'aile et est la longueur de l'aile, y compris le bout de l'aile.







En plus du nombre de Reynolds, il existe au moins deux autres paramètres sans dimension pertinents. Une aile a trois échelles de vitesse : la vitesse de battement par rapport au corps ( u ), la vitesse vers l'avant du corps ( U 0 ) et la vitesse de tangage (Ω c ). Leurs rapports forment deux variables sans dimension, U 0 / u et c/ u , la première est souvent appelée rapport d'avance, et elle est également liée à la fréquence réduite, fc / U 0 .
Si une aile d'insecte est rigide, par exemple, une aile de drosophile l' est approximativement, son mouvement par rapport à un corps fixe peut être décrit par trois variables : la position de la pointe en coordonnées sphériques , (Θ(t),Φ(t) ), et l' angle de tangage ψ(t), autour de l'axe reliant la racine et la pointe. Pour estimer les forces aérodynamiques à partir d'une analyse pale-élément, il est également nécessaire de déterminer l' angle d'attaque (α). L'angle d'attaque typique à 70 % d'envergure varie de 25° à 45° chez les insectes en vol stationnaire (15° chez les colibris). Malgré la richesse des données disponibles pour de nombreux insectes, relativement peu d'expériences rapportent la variation temporelle de au cours d'un AVC. Parmi ceux-ci figurent des expériences en soufflerie d'un criquet attaché et d'une mouche attachée, et le vol stationnaire libre d'une mouche des fruits.
Parce qu'elles sont relativement faciles à mesurer, les trajectoires de bout d'aile ont été rapportées plus fréquemment. Par exemple, en sélectionnant uniquement des séquences de vol qui ont produit suffisamment de portance pour supporter un poids, vous montrerez que le bout de l'aile suit une forme elliptique. Des formes non croisées ont également été signalées pour d'autres insectes. Indépendamment de leurs formes exactes, le mouvement de blocage indique que les insectes peuvent utiliser la traînée aérodynamique en plus de la portance pour supporter leur poids.
En vol stationnaire
| Paramètres de vol | Vitesse (m/s) | Battements/s |
|---|---|---|
| Libellule Aeshnide | 7.0 | 38 |
| frelon | 5.7 | 100 |
| Sphinx colibri | 5.0 | 85 |
| Taon | 3.9 | 96 |
| Hoverfly | 3.5 | 120 |
| Bourdon | 2.9 | 130 |
| Abeille | 2.5 | 250 |
| Mouche domestique | 2.0 | 190 |
| Demoiselle | 1.5 | 16 |
| Scorpionfly | 0,49 | 28 |
| Grand papillon blanc | 2.5 | 12 |
| Thrips (clap and fling) | 0,3 | 254 |
.jpg)
De nombreux insectes peuvent planer ou rester au même endroit dans les airs, en battant rapidement des ailes. Cela nécessite une stabilisation latérale ainsi que la production de portance. La force de levage est principalement produite par la course descendante. Lorsque les ailes poussent vers le bas sur l'air environnant, la force de réaction résultante de l'air sur les ailes pousse l'insecte vers le haut. Les ailes de la plupart des insectes sont évoluées de sorte que, pendant la course ascendante, la force sur l'aile est faible. Étant donné que le coup bas et le coup de retour forcent l'insecte vers le haut et vers le bas respectivement, l'insecte oscille et s'enroule en restant dans la même position.
La distance à laquelle l'insecte tombe entre les battements d'ailes dépend de la vitesse à laquelle ses ailes battent : plus il bat lentement, plus l'intervalle de chute est long et plus il tombe loin entre chaque battement d'aile. On peut calculer la fréquence des battements d'ailes nécessaire à l'insecte pour maintenir une stabilité donnée dans son amplitude. Pour simplifier les calculs, il faut supposer que la force de portance est à une valeur constante finie pendant que les ailes descendent et qu'elle est nulle pendant que les ailes montent. Pendant l'intervalle de temps Δ t du battement d'aile ascendant, l'insecte descend d'une distance h sous l'influence de la gravité.

La course vers le haut restaure ensuite l'insecte dans sa position d'origine. Typiquement, il peut être nécessaire que la position verticale de l'insecte ne change pas de plus de 0,1 mm (c'est-à-dire, h = 0,1 mm). Le temps maximum autorisé pour la chute libre est alors

Étant donné que les mouvements de montée et de descente des ailes sont à peu près de durée égale, la période T pour une aile complète de va-et-vient est de deux fois Δ r , c'est-à-dire

La fréquence des battements, f, c'est-à-dire le nombre de battements d'ailes par seconde, est représentée par l'équation :

Dans les exemples utilisés, la fréquence utilisée est de 110 battements/s, qui est la fréquence typique trouvée chez les insectes. Les papillons ont une fréquence beaucoup plus lente avec environ 10 battements/s, ce qui signifie qu'ils ne peuvent pas planer. D'autres insectes peuvent être capables de produire une fréquence de 1000 battements/s. Pour remettre l'insecte dans sa position verticale d'origine, la force ascendante moyenne pendant la course descendante, F av , doit être égale à deux fois le poids de l'insecte. Notez que puisque la force ascendante sur le corps de l'insecte n'est appliquée que pendant la moitié du temps, la force ascendante moyenne sur l'insecte est simplement son poids.
Entrée de puissance
On peut maintenant calculer la puissance nécessaire pour maintenir le vol stationnaire en considérant à nouveau un insecte de masse m 0,1 g, la force moyenne, F av , appliquée par les deux ailes lors de la course descendante est de deux fois le poids. Parce que la pression appliquée par les ailes est uniformément répartie sur toute la surface de l'aile, cela signifie que l'on peut supposer que la force générée par chaque aile agit à travers un seul point à la section médiane des ailes. Lors de la course descendante, le centre des ailes parcourt une distance verticale d . Le travail total effectué par l'insecte au cours de chaque course descendante est le produit de la force et de la distance ; C'est,

Si les ailes oscillent dans le battement à un angle de 70°, alors dans le cas présenté pour l'insecte avec des ailes de 1 cm de long, d est de 0,57 cm. Par conséquent, le travail effectué à chaque coup par les deux ailes est :

L'énergie est utilisée pour élever l'insecte contre la gravité. L'énergie E nécessaire pour soulever la masse de l'insecte de 0,1 mm à chaque descente est :

Il s'agit d'une fraction négligeable de l'énergie totale dépensée qui, de toute évidence, est la plus grande partie de l'énergie dépensée dans d'autres processus. Une analyse plus détaillée du problème montre que le travail effectué par les ailes est converti principalement en énergie cinétique de l'air qui est accélérée par la course descendante des ailes. La puissance est la quantité de travail effectué en 1 s ; dans l'insecte utilisé comme exemple, fait 110 coups vers le bas par seconde. Par conséquent, sa puissance de sortie P est de coups par seconde, ce qui signifie que sa puissance de sortie P est :

Puissance de sortie
Dans le calcul de la puissance utilisée en vol stationnaire, les exemples utilisés ont négligé l'énergie cinétique des ailes en mouvement. Les ailes des insectes, si légères soient-elles, ont une masse finie ; par conséquent, lorsqu'ils se déplacent, ils possèdent de l'énergie cinétique. Étant donné que les ailes sont en mouvement de rotation, l'énergie cinétique maximale pendant chaque coup d'aile est :

Ici I est le moment d' inertie de l'aile et max est la vitesse angulaire maximale lors de la course de l'aile. Pour obtenir le moment d'inertie de l'aile, nous supposerons que l'aile peut être approchée par une tige mince pivotée à une extrémité. Le moment d'inertie de l'aile est alors :

Où l est la longueur de l'aile (1 cm) et m est la masse de deux ailes, qui peut être typiquement de 10 -3 g. La vitesse angulaire maximale, ω max , peut être calculée à partir du maximum linéaire vitesse , ν max , au centre de l'aile:

Au cours de chaque course du centre de l'aile se déplace avec une vitesse linéaire moyenne ν av donnée par la distance d parcourue par le centre de l'aile divisée par la durée At de la course de l' aile. D'après notre exemple précédent, d = 0,57 cm et t = 4,5×10 −3 s. Par conséquent:

La vitesse des ailes est nulle au début et à la fin de la course de l'aile, ce qui signifie que la vitesse linéaire maximale est supérieure à la vitesse moyenne. Si nous supposons que la vitesse oscille ( sinusoïdalement ) le long de la trajectoire de l'aile, la vitesse maximale est deux fois plus élevée que la vitesse moyenne. La vitesse angulaire maximale est donc :

Et l'énergie cinétique est donc :

Puisqu'il y a deux coups d'aile (la course ascendante et la course descendante) dans chaque cycle du mouvement de l'aile, l'énergie cinétique est de 2 × 43 = 86 erg . C'est à peu près autant d'énergie que le vol stationnaire lui-même consomme.
Élasticité

Les insectes gagnent de l'énergie cinétique, fournie par les muscles, lorsque les ailes accélèrent . Lorsque les ailes commencent à décélérer vers la fin de la course, cette énergie doit se dissiper. Lors de la descente, l'énergie cinétique est dissipée par les muscles eux-mêmes et convertie en chaleur (cette chaleur est parfois utilisée pour maintenir la température corporelle centrale). Certains insectes sont capables d'utiliser l'énergie cinétique du mouvement ascendant des ailes pour faciliter leur vol. Les articulations des ailes de ces insectes contiennent un coussin de protéine élastique, semblable à du caoutchouc, appelé résiline . Lors de la remontée de l'aile, la résine est étirée. L'énergie cinétique de l'aile est convertie en énergie potentielle dans la résine étirée, qui stocke l'énergie un peu comme un ressort. Lorsque l'aile descend, cette énergie est libérée et aide à la descente.
En utilisant quelques hypothèses simplificatrices, nous pouvons calculer la quantité d'énergie stockée dans la résine étirée. Bien que la résine soit pliée en une forme complexe, l'exemple donné montre le calcul sous la forme d'une tige droite de surface A et de longueur. De plus, nous supposerons que tout au long de l'étirement, la résine obéit à la loi de Hooke . Ceci n'est pas strictement vrai car la résiline est étirée d'une quantité considérable et donc à la fois l'aire et le module de Young changent au cours du processus d'étirement. L'énergie potentielle U stockée dans la résine étirée est :

Ici E est le module de Young pour résiline, qui a été mesurée à 1,8 x 10 7 dyn / cm 2 . Typiquement chez un insecte de la taille d'une abeille, le volume de la résiline peut être équivalent à un cylindre de 2x10 -2 cm de long et de 4x10 -4 cm 2 de surface. Dans l'exemple donné, la longueur de la tige de résine est augmentée de 50 % lorsqu'elle est étirée. C'est-à-dire que est de 10 -2 cm. Par conséquent, dans ce cas, l'énergie potentielle stockée dans la résine de chaque aile est :

L'énergie stockée dans les deux ailes pour un insecte de la taille d'une abeille est de 36 erg, ce qui est comparable à l'énergie cinétique de la course ascendante des ailes. Les expériences montrent que jusqu'à 80 % de l'énergie cinétique de l'aile peut être stockée dans la résine.
Attelage d'aile

Certains ordres d'insectes à quatre ailes, tels que les lépidoptères, ont développé des mécanismes de couplage morphologique des ailes dans l' imago qui rendent ces taxons fonctionnellement à deux ailes. Toutes les formes sauf les plus basales présentent ce couplage alaire.
Les mécanismes sont de trois types différents - jugal, frénulo-rétinaculaire et amplexiforme :
- Les groupes les plus primitifs ont une zone en forme de lobe élargie près de la marge postérieure basale, c'est-à-dire à la base de l'aile antérieure, un jugum , qui se replie sous l'aile postérieure en vol.
- D'autres groupes ont un frein sur l'aile postérieure qui s'accroche sous un rétinaculum sur l'aile antérieure.
- Chez presque tous les papillons et chez les Bombycoidea (sauf les Sphingidae ), il n'y a pas d'arrangement de frein et de rétinaculum pour coupler les ailes. Au lieu de cela, une zone humérale élargie de l'aile postérieure est largement recouverte par l'aile antérieure. Malgré l'absence de liaison mécanique spécifique, les ailes se chevauchent et fonctionnent en phase. Le coup de force de l'aile antérieure abaisse l'aile postérieure à l'unisson. Ce type de couplage est une variante de type frénate mais où le frein et le rétinaculum sont complètement perdus.
Biochimie
La biochimie du vol des insectes a fait l'objet d'études considérables. Alors que de nombreux insectes utilisent des glucides et des lipides comme source d'énergie pour voler, de nombreux coléoptères et mouches utilisent l' acide aminé proline comme source d'énergie. Certaines espèces utilisent également une combinaison de sources et les mites telles que Manduca sexta utilisent des glucides pour l'échauffement avant le vol.
Évolution et adaptation

Au cours de la période carbonifère , il y a 350 à 400 millions d'années, alors qu'il n'y avait que deux grandes masses continentales, les insectes ont commencé à voler. Parmi les plus anciens fossiles d'insectes ailés se trouve Delitzschala , un paléodictyoptère du Carbonifère inférieur ; Rhyniognatha est plus ancien, du Dévonien inférieur , mais on ne sait pas s'il avait des ailes, ou s'il s'agissait bien d'un insecte.
Comment et pourquoi les ailes d'insectes se sont développées n'est pas bien compris, en grande partie en raison de la rareté des fossiles appropriés de la période de leur développement dans le Carbonifère inférieur. Il y a eu historiquement trois théories principales sur les origines du vol des insectes. La première était qu'il s'agissait de modifications des branchies abdominales mobiles , comme on en trouve sur les naïades aquatiques des éphémères . L'analyse phylogénomique suggère que les polynéoptères , le groupe d'insectes ailés qui comprend les sauterelles, ont évolué à partir d'un ancêtre terrestre, ce qui rend improbable l'évolution des ailes à partir des branchies. Une étude supplémentaire du comportement de saut des larves d'éphémères a déterminé que les branchies trachéales ne jouent aucun rôle dans le guidage de la descente des insectes, fournissant une preuve supplémentaire contre cette hypothèse évolutive. Cela laisse deux théories historiques majeures : que les ailes se sont développées à partir de lobes paranotaux, prolongements du terga thoracique ; ou qu'ils provenaient de modifications de segments de jambe, qui contenaient déjà des muscles.
Hypothèse épicoxale (ouïe abdominale)
.JPG)
De nombreux entomologistes, dont Landois en 1871, Lubbock en 1873, Graber en 1877 et Osborn en 1905, ont suggéré qu'une origine possible des ailes d'insectes pourrait avoir été les branchies abdominales mobiles trouvées chez de nombreux insectes aquatiques, comme sur les naïades des éphémères. Selon cette théorie, ces branchies trachéales , qui ont commencé leur chemin en tant que sorties du système respiratoire et au fil du temps ont été modifiées en fonctions locomotrices, se sont finalement développées en ailes. Les branchies trachéales sont équipées de petites ailettes qui vibrent perpétuellement et ont leurs propres petits muscles droits.
Hypothèse paranotale (tergal)
L'hypothèse du lobe paranotal ou tergal (paroi dorsale du corps), proposée par Fritz Müller en 1875 et retravaillée par G. Crampton en 1916, Jarmila Kulakova-Peck en 1978 et Alexander P. Rasnitsyn en 1981 entre autres, suggère que les ailes de l'insecte se sont développées à partir de lobes paranotaux, une préadaptation trouvée dans les fossiles d' insectes qui aurait aidé à la stabilisation en sautant ou en tombant. En faveur de cette hypothèse, la tendance de la plupart des insectes, lorsqu'ils sont surpris en grimpant sur des branches, à s'échapper en tombant au sol. De tels lobes auraient servi de parachutes et auraient permis à l'insecte de se poser plus doucement. La théorie suggère que ces lobes se sont progressivement agrandis et qu'à un stade ultérieur, ils ont développé une articulation avec le thorax. Plus tard encore apparaîtraient les muscles pour déplacer ces ailes grossières. Ce modèle implique une augmentation progressive de l'efficacité des ailes, en commençant par le parachutisme , puis le vol plané et enfin le vol actif . Pourtant, le manque de preuves fossiles substantielles du développement des articulations et des muscles des ailes pose une difficulté majeure à la théorie, tout comme le développement apparemment spontané de l'articulation et de la nervation, et il a été largement rejeté par les experts dans le domaine.
Hypothèse d'endite-sortie (pleurale)

En 1990, JWH Trueman a proposé que l'aile soit adaptée à partir d' endites et d'exites , des appendices situés sur les aspects internes et externes respectifs du membre primitif de l' arthropode , également appelée hypothèse pleurale. Ceci était basé sur une étude de Goldschmidt en 1945 sur Drosophila melanogaster , dans laquelle une variation appelée "pod" (pour podomeres, segments de membres) présentait une mutation qui transformait les ailes normales. Le résultat a été interprété comme un arrangement de jambe à triple articulation avec quelques appendices supplémentaires mais sans le tarse, là où se trouverait normalement la surface costale de l'aile. Cette mutation a été réinterprétée comme une preuve solide d'une fusion de l'exite dorsale et de l'endite, plutôt que d'une jambe, les appendices s'accordant beaucoup mieux avec cette hypothèse. L'innervation, l'articulation et la musculature nécessaires à l'évolution des ailes sont déjà présentes dans les segments des membres.
Autres hypothèses
D'autres hypothèses incluent la suggestion de Vincent Wigglesworth de 1973 selon laquelle les ailes se sont développées à partir de protubérances thoraciques utilisées comme radiateurs .
Adrian Thomas et Åke Norberg ont suggéré en 2003 que les ailes pouvaient avoir évolué initialement pour naviguer à la surface de l'eau, comme on le voit chez certains phlébotomes .
Stephen P. Yanoviak et ses collègues ont proposé en 2009 que l'aile dérive d'une descente en plané aérien dirigé - un phénomène de vol en amont trouvé chez certains aptérygotes , un taxon frère sans ailes des insectes ailés.
Double origine
Des biologistes comme Averof, Niwa, Elias-Neto et leurs collègues ont commencé à explorer l'origine de l'aile d'insecte en utilisant evo-devo en plus des preuves paléontologiques. Cela suggère que les ailes sont homologues en série avec les structures tergale et pleurale, résolvant potentiellement le débat vieux de plusieurs siècles. Jakub Prokop et ses collègues ont trouvé en 2017 des preuves paléontologiques des coussinets d'ailes nymphales paléozoïques que les ailes avaient en effet une si double origine.
Voir également
- Vol d'oiseau
- Animaux volants et planeurs
- Fourmi planante
- Migration des insectes
- Véhicules aériens sans pilote
Les références
Lectures complémentaires
- Dickinson, MH ; Lehmann, FO ; Sané, SP (1999). « La rotation des ailes et la base aérodynamique du vol des insectes ». Sciences . 284 (5422) : 1954-1960. doi : 10.1126/science.284.5422.1954 . PMID 10373107 .
- Ellington, Charles P.; Van Den Berg, Coen ; Willmott, Alexander P.; Thomas, Adrien LR (1996). « Les tourbillons de pointe dans le vol des insectes ». Nature . 384 (6610) : 626-630. Bibcode : 1996Natur.384..626E . doi : 10.1038/384626a0 . S2CID 4358428 .
- Ellington, CP (1984). "L'aérodynamique du vol des insectes en vol stationnaire. I. L'analyse quasi-stable" . Transactions philosophiques de la Royal Society B : Sciences biologiques . 305 (1122) : 1-15. Bibcode : 1984RSPTB.305 .... 1E . doi : 10.1098/rstb.1984.0049 .
- Ellington, CP (1999). « La nouvelle aérodynamique du vol des insectes : applications aux véhicules micro-aériens » . Le Journal de biologie expérimentale . 202 (Pt 23) : 3439-3448. doi : 10.1242/jeb.202.23.3439 . PMID 10562527 .
- Lewin, GC ; Haj-Hariri, H. (2003). « Modélisation de la génération de poussée d'un profil aérodynamique bidimensionnel dans un écoulement visqueux ». Journal de mécanique des fluides . 492 : 339-362. Bibcode : 2003JFM ... 492..339L . doi : 10.1017/S0022112003005743 . S2CID 122077834 .
- Pringle, JWS (1975). Vol d'insectes . Lecteurs de biologie d'Oxford. 52 .
- Sané, SP (2003). " L'aérodynamique du vol des insectes " . Le Journal de biologie expérimentale . 206 (Pt 23) : 4191-208. doi : 10.1242/jeb.00663 . PMID 14581590 .
- Sauvage, SB ; Newman, BG ; Wong, DTM (1979). "Le rôle des tourbillons et des effets instables pendant le vol stationnaire des libellules" . Le Journal de biologie expérimentale . 83 (1) : 59-77. doi : 10.1242/jeb.83.1.59 .
- Smyth, T., Jr. (1985). "Systèmes musculaires". Dans Blum, MS Blum (éd.). Fondements de la physiologie des insectes . John Wiley et fils. p. 227-286.
- Sotavalta, O. (1953). « Enregistrements de la fréquence élevée des coups d'aile et des vibrations thoraciques chez certains moucherons » . Biol. Taureau. Trou des bois . 104 (3) : 439-444. doi : 10.2307/1538496 . JSTOR 1538496 .
- Van Den Berg, C. ; Ellington, CP (1997). "Le sillage tourbillonnaire d'un sphinx modèle 'planant'" . Transactions philosophiques de la Royal Society B : Sciences biologiques . 352 (1351) : 317-328. Bibcode : 1997RSPTB.352..317V . doi : 10.1098/rstb.1997.0023 . PMC 1691928 .
- Walker, JA (septembre 2002). « Élévateur rotatif : quelque chose de différent ou plus ou moins la même chose ? » . Le Journal de biologie expérimentale . 205 (Pt 24) : 3783-3792. doi : 10.1242/jeb.205.24.3783 . PMID 12432002 .
- Zbikowski, R. (2002). « Sur la modélisation aérodynamique d'une aile battante semblable à un insecte en vol stationnaire pour les véhicules micro aériens ». Transactions philosophiques de la Royal Society A: Sciences mathématiques, physiques et techniques . 360 (1791) : 273-290. Bibcode : 2002RSPTA.360..273Z . doi : 10.1098/rsta.2001.0930 . PMID 16210181 . S2CID 2430367 .
Liens externes
- Le rôle d'un insecte dans le développement des micro-véhicules aériens
- Micro-véhicules aériens à ailes battantes ressemblant à des insectes
- La nouvelle aérodynamique du vol des insectes : applications aux véhicules micro-aériens
- L'aérodynamique du vol des insectes
- Muscles de vol
- Visualisation du flux des mécanismes aérodynamiques des papillons
- Le vol du bourdon
- Vol d'insectes
{kind=link}